|
|
Журнал "Вопросы современной альгологии" (переход на сайт algology.ru) |
Семинары и Конференции (переход на сайт algology.ru) |
Партнеры (переход на сайт algology.ru) | Контакты | ENGLISH |
|
|
Флуоресценция отдельных клеток водоросли Conticribra weissflogii при гиперосмотическом стрессе
Ильяш Л.В., Курочкина В.А., Белевич Т.А., Погосян С.И. Ludmila V. Il’yash, Viktoria A. Kurochkina, Tatiana A. Belevich,
Московский Государственный Университет имени М.В.Ломоносова,
УДК 581.132
У диатомовой водоросли Conticribra weissflogii, акклимированной к солености 17,5‰, прослежена динамика относительной переменной флуоресценции отдельных клеток (Fvи/Fmи) и удельной (на клетку) фотосинтетической фиксации углерода после пересева на среды с соленостью 35‰. Выявлена более высокая степень изменчивости Fvи/Fmи в условиях осмотического стресса. Ключевые слова: флуоресценция; популяционная гетерогенность; осмотический стресс; Conticribra weissflogii.
Стресс определяют как изменение условий роста, ведущее к нарушению гомеостаза клеточного метаболизма и вызывающее перестройку процессов метаболизма, называемую акклимацией (Shulaev et al., 2008). У фотоавтотрофных организмов наиболее чувствительным сенсором стресса выступают хлоропласты. Одним из показателей состояния фотосинтетического аппарата является потенциальная квантовая эффективность фотосистемы 2, количественно характеризуемая величиной относительной переменной флуоресценции (Fv/Fm). Способность популяции акклимироваться к стрессу и, в конечном счете, выживать в изменяющихся экологических условиях в значительной мере определяется внутрипопуляционной гетерогенностью, т.е. различием особей в пределах одной популяции. И если внутрипопуляционной изменчивости животных и высших растений посвящено значительное число работ, то сведения о гетерогенности популяций микроскопических водорослей крайне малочислены. Лишь в последнее десятилетие с развитием новых методов, позволяющих оценивать те или иные параметры отдельных клеток, появились данные о вариабельности отклика отдельных клеток на изменение уровня контролирующего фактора и лимитирующего ресурса (Воронова и др., 2009; Peloquin, Smith, 2006; Heraud et al., 2008). Соленость является одним из основных факторов, контролирующих фотосинтетическую активность микроводорослей, особенно в эстуарных экосистемах и ледовых биотопах. Как снижение солености среды по сравнению с оптимальной (гипоосмотический стресс), так и увеличение (гиперосмотический стресс) вызывают изменение скорости роста популяции (Rijstenbil et al., 1989a, b; Cifuentes et al., 2001), скорости фотосинтетической фиксации углерода (Радченко, Ильяш, 2006), содержания фотосинтетических пигментов (Cifuentes et al., 2001), скорости потребления биогенных элементов (Rijstenbil et al., 1989 a, b). Все эти параметры представляют собой усредненные значение для популяции, тогда как сведения по изменчивости функциональных параметров отдельных клеток в популяции в ответ на осмотический стресс отсутствуют. В настоящей работе представлены данные о вариабельности относительной переменной флуоресценции отдельных клеток в популяции диатомовой водоросли Conticribra weissflogii при акклимации к гиперосмотическому стрессу Материалы и методы исследования Материалом для исследования послужила альгологически чистая культуры диатомовой водорослий Conticribra weissflogii (Grunow) Stachura-Suchoples et Williams (синоним Thalassiosira weissflogii Grunow). Эта водоросль отмечается как в морских (Hasle, Syvertsen, 1997), так и в пресных водах (Генкал, Михайловский, 2009), и отнесена к солоновато-водным видам (Stachura-Suphoples, Williams, 2009). В течение нескольких месяцев водоросли акклимировали к солености 17,5‰ путем многократных пересевов на среду, приготовленной на основе искусственной морской воды (сoлевaя смесь фиpмы «SЕRA») и обогащённой биогенными элементами по прописи среды f/2 (Guillаrd, Rythеr, 1962). Схема экспериментов. Водоросли, акклимированные к солености 17,5‰ из культуры, находящейся на экспоненциальной стадии роста, отделяли от среды центрифугированием (5 тыс. оборотов/мин) и высевали на среду с соленостью 17,5‰ (контроль) и 35‰ (гиперосмотические условия). При этом добавки биогенных элементов не вносили. Водоросли росли в накопительном режиме при температуре 20°С, освещенности 75 мкЕ/м2∙с и продолжительности светового периода 14 ч. Культивирование на каждой солености проводили в 300 мл среды в двух повторностях. Оцениваемые параметры. Численность водорослей (N) определяли методом прямого счета в камере Горяева под световым микроскопом. Для каждого варианта культур просчитывали не менее трех камер. Сразу после посева водорослей на среды с разной соленостью, а также через 24 ч и 48 ч оценивали фотосинтетическую фиксацию углерода скляночным методом в радиоуглеродной модификации (Steemann Nielsen, 1952). Время экспозиции составляло 4 ч, объем экспонируемых проб – 10 мл. Через 2, 4, 6, 24 и 48 ч после посева водорослей на среды с разной соленостью определяли параметры флуоресценции хлорофилла у водорослей, выдержанных 15 минут в темноте. У отдельных клеток водорослей при помощи переоборудованного флуоресцентного микроскопа ЛЮМАМ И3 по ранее описанной методике (Воронова и др., 2009) оценивали флуоресценцию при открытых (Foи) и закрытых (Fmи) реакционных центрах и рассчитывали относительную переменную флуоресценцию Fvи/Fmи =(Fmи-Foи)/Fmи. Время измерения Foи и Fmи составляло около 10 с. При каждом измерении выборка составляла 100 клеток. Величины Fvи/Fmи изменяются от 0 до 0,72 (Falkowski, Raven, 1997). Высокие значения характерны для водорослей в хорошем физиологическом состоянии, тогда как низкие свидетельствуют о состоянии стресса. Клетки с Fvи/Fmи < 0,1 являются фотосинтетически не активными и нежизнеспособными (Franklin et al. 2009; Reeves et al., 2011). Учитывая опубликованные данные (Воронова и др., 2009; Franklin et al., 2009; Reeves et al., 2011) далее по тексту принимали следующее ранжирование фотосинтетической активности по величинам Fvи/Fmи < 0,1 - фотосинтетически не активные клетки; 0.1–0,29 – низкая; 0,30–0,49 – средняя; 0,50–0,59 – высокая; ≥ 0,60 – максимальная фотосинтетическая активность. Статистический анализ данных. Для сравнения степени варьирования Fvи/Fmи во времени и в популяциях, росших при разной солености, использовали коэффициент вариации CV. Достоверность различий CV (Pcv) оценивали по критерию Флайгнера-Киллена (Fligner-Killeen Test), а достоверность различий средних значений Fvи/Fmи (Рср) – по критерию Манна-Уитни с использованием программы PAST (PAleontological STatistics) Version 2.04 (http://folk.uio.no/ohammer/past/). Результаты После посева водорослей на среды, не содержащие биогенные элементы, численность водорослей увеличивалась (таб.1). Размножение клеток обеспечивалось, по-видимому, внутриклеточными запасами биогенных элементов. В контрольной культуре рост продолжался сутки, в условиях гиперосмотического стресса - двое суток. Скорость роста в первые сутки была выше в контроле. Удельная (на клетку) фотосинтетическая фиксация углерода (P/N) в первые 4 ч после пересева в контрольной культуре и при 35 ‰ не различалась, однако через сутки P/N стала ниже в контроле. Это обусловлено достижением контрольной культуры максимальной численности, и исчерпанием внутриклеточного запаса биогенных элементов.
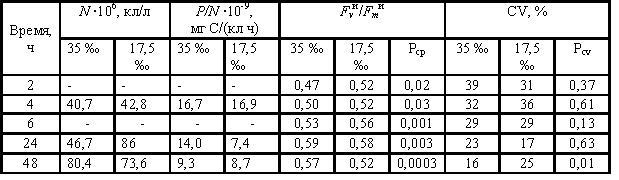
Таблица 1. Численность (N), удельная (на клетку) фотосинтетическая фиксация углерода (P/N), средние значения относительной переменной флуоресценции отдельных клеток (Fvи/Fmи), коэффициент вариации Fvи/Fmи (CV), достоверность различий средних значений Fvи/Fmи (Рср) и CV (Pcv) в культурах Conticribra weissflogii, росших при солености 35 ‰ и 17,5 ‰.
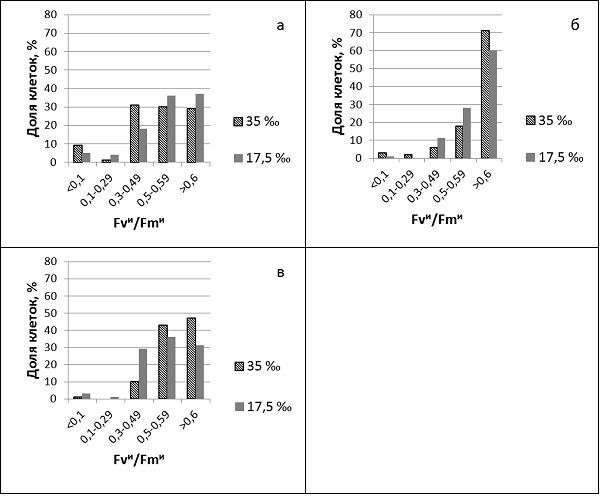
Вариабельность клеток C. weissflogii по Fvи/Fmи отмечена как в контрольной культуре, так и у водорослей, росших в условиях гиперосмотических условиях (рис.1). В контроле средние значения Fvи/Fmи достоверно возросли через 6 ч роста (Рср = 0,003). Нежизнеспособные клетки отмечены через 2 ч после пересева, по мере роста их доля снижалась, а доля клеток с максимальной фотосинтетической активностью возрастала. У водорослей, перенесенных в гиперосмотические условия значения Fvи/Fmи снизились, особенно в первые 2 ч. Средние значения Fvи/Fmи были достоверно ниже контроля в течение, по крайней мере, шести часов. Уже в первые 2 ч возросла доля нежизнеспособных клеток и клеток со средней фотосинтетической активностью (рис.1). При разном распределении клеток по Fvи/Fmи в популяциях, росших при 35‰ и 17,5‰, степень варьирования Fvи/Fmи достоверно не различалась. Степень варьирования Fvи/Fmи в гиперосмотических условиях через 6 ч достоверно снизилась (Рcv=0,009). Более высокие средние значения Fvи/Fmи через 24 ч в гиперосмотических условиях обусловлены более высокой по сравнению с контролем долей клеток с максимальной фотосинтетической активностью.
Рис.1. Доля клеток (%) с разной относительной переменной флуоресценцией (Fvи/Fmи) в культурах, росших 2 ч (а), 24 ч (б) и 48 ч (в) при солености 35 ‰ и 17,5 ‰.
Обсуждение результатов Гетерогенность популяций C. weissflogii по Fvи/Fmи выявлена как в условиях осмотического стресса, так и в его отсутствие. Обращает на себя внимание присутствие нежизнеспособных клеток в контрольной популяции, не испытывающей осмотический стресс. Доля нежизнеспособных клеток не превышала 5%, что соответствует доли мертвых клеток в активно растущих культурах других водорослей (Veldhuis et al., 2001). При снижении численности контрольной популяции на вторые сутки доля нежизнеспособных клеток увеличивалась. В условиях гиперосмотического стресса доля нежизнеспособных клеток возрастала уже в первые 2 ч, доля клеток с высокой и максимальной фотосинтетической активностью снижалась, что привело к снижению средних для популяции значений Fvи/Fmи. Все это свидетельствует о нарушениях в работе фотосинтетического аппарата. Гиперосмотический стресс вызывает у водорослей повышенное образование активных форм кислорода (АФК) таких как H2O2, O2-, 1О2, ОН-, которое уже не может быть нейтрализовано антиоксидантной клеточной системой (Rijstenbil, 2005). Избыточное количество АФК окисляет пигменты, белки, а также тилакоидные мембраны, что ведет к снижению фотосинтетической активности (Foyer et al., 1994; Lu, Vonshak, 1999). Помимо этого АФК окисляют липиды, что обусловливает увеличение проницаемости клеточных мембран (Rijstenbil, 2005). Нарушение проницаемости мембран является первой стадией в процессе отмирания водорослей, за ней следуют деградация фотосинтетических пигментов, фрагментация ядерной ДНК и лизис клеток (Veldhuis et al., 2001). Обращает на себя внимание тот факт, что в первые часы после перенесения водорослей в гиперосмотические условия, регистрируется снижение потенциальной квантовой эффективности фотосистемы 2, тогда как скорость фотосинтетической фиксации углерода остается без изменения. Это свидетельствует о более высокой чувствительности процессов световой стадии фотосинтеза к стрессовым условиям и обусловливает перспективность оценки параметров флуоресценции отдельных клеток для выявления неблагоприятных для водорослей условий. Через сутки Conticribra weissflogii акклимируется к двухкратному увеличению солености. Доля нежизнеспособных клеток снижается, а доля клеток с максимальной активностью увеличивается. Средние значения Fvи/Fmи и удельная фотосинтетическая фиксация углерода становятся достоверно выше, чем в контрольной популяции, которая в этот период уже испытывает клеточный дефицит биогенных элементов. Гетерогенность популяций по Fvи/Fmи – от фотосинтетически неактивных клеток до клеток с максимальной фотосинтетической активностью отмечается как в условиях осмотического стресса, так и в его отсутствие. Такая популяционная гетерогенность физиологического состояния может быть обусловлена генотипической неоднородностью популяции. Популяциям планктонных водорослей присуща значительная генотипическая неоднородность клеток (Rynearson, Armbrust, 2004, 2005; Iglesias-Rodriguez et al., 2006; Dassow von et al., 2008). Внутрипопуляционную вариабельность таких параметров как размеры клеток, скорость роста (Rynearson, Armbrust, 2004), устойчивость к токсикантам (Lopez-Rodas et al., 2001) связывают именно с различием клеток по генотипу. Не следует также исключать, что вариабельность Fvи/Fmи в условиях осмотического стресса может быть обусловлена также гетерогенностью популяции по экспрессии генов в отдельных клетках (Elowitz et al., 2002). В результате генотипической неоднородности популяции часть клеток оказывается более чувствительной к изменению солености среды, и нарушения в работе их фотосинтетического аппарата сказываются в большей степени. Также генотипическая неоднородность ведет к тому, что клетки в пределах одной популяции различаются по способности акклимироваться к осмотическому стрессу – одни клетки акклимируются быстрее, другие – медленнее. В процессе акклимации вариабельность клеток по Fvи/Fmи снижается, т.е. гетерогенность популяции в большей степени проявляется в условиях стресса.
Список литературы 1. Воронова Е. Н., Ильяш Л. В., Погосян С. И., Уланова А. Ю., Маторин Д. Н., Ман-ги Хо, Рубин А. Б. Внутрипопуляционная гетерогенность параметров флуоресценции у морской планктонной водоросли Thalassiosira weissflogii при разной обеспеченности азотом // Микробиология. 2009. Т. 78. № 4. С. 469–478. 2. Генкал С.И., Куликовский М.С. К морфологии, экологии и распространению Thalassiosira weissflogii (Bacillariophyta) // Поволжский экологический журнал. 2009. № 3. С. 183–189. 3. Радченко И.Г., Ильяш Л.В. Рост и фотосинтетическая активность диатомовой водоросли Thalassiosira weissflogii при снижении солености // Известия АН. Серия Биологическая. 2006. № 3. С. 306-313. 4. Cifuentes A.S., Gonza´lez M.A., Inostroza I., Aguilera A. Reappraisal of physiological attributes of nine strains of Dunaliella (Chlorophyceae): growth and pigment content across a salinity gradient a // J. Phycol. 2001. V. 37. P. 334–344. 5. Dassow von P., Petersen T.W., Chepurnov V. A., Armbrust E.V. Inter- and intraspecific relationships between nuclear DNA content and cell size in selected members of the centric diatom genus Thalassiosira (Bacillariophyceae) // J. Phycology. 2008. V. 44. P. 335–349. 6. Elowitz M., Levine A., Siggia E., Swain P. Stochastic gene expression in a single cell // Science. 2002. V. 297. P. 1183–1186. 7. Falkowski P.G., Raven J.A. Aquatic photosynthesis. Malden, Massachusetts: Blackwell Science, 1997. 375 p. 8. Foyer C.H., Lelandais M., Kunert K.J. Photooxidative stress in plants // Physiol. Plant. 1994. V. 92. P. 696–717. 9. Franklin D.J., Choi C.J., Hughes C., Malin G., Berges J.A. Effect of dead phytoplankton cells on the apparent efficiency of photosystem II // Mar. Ecol. Prog. Ser. 2009. V. 382. P. 35–40. 10. Guillard R.R.L., Ryther J.H. Studies on marine diatoms. I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. // Can. J. Microbiol. 1962. V.8. P. 229-239. 11. Hasle G.R., Syvertsen E. E. Marine diatoms / Identifying Marine Phytoplankton. Ed. C.R. Tomas. San Diego, California: Academic Press, 1997. P.5–385. 12. Heraud P., Stojkovic S., Beardall J., McNaughton D., Wood B.R. Intercolonial variability in macromolecular composition in P-starved and P-replete Scenedesmus population revealed by infrared microspectroscopy // J. Phycol. 2008. V. 44. P. 1335–1339. 13. http://folk.uio.no/ohammer/past/ - программа PAST (PAleontological STatistics), University of Oslo (дата обращения 10.10.2012). 13. Iglesias-Rodriguez M.D., Schofield O.M., Batley J., Medlin L.K., Hayes P.K. Intraspecific genetic diversity in the marine coccolithophore Emiliania huxleyi (Prymnesiophyceae): the use of microsatellite analysis in marine phytoplankton population studies // J. Phycol. 2006. V. 42. P. 526-536. 14. Lopez-Rodas V., Agrelo M., Carrillo E., Ferrero L., Larrauri A., Martin-Otero L., Costas E. Resistance of microalgae to modern water contaminants as the result of rare spontaneous mutations // Eur. J. Phycology. 2001. V. 36. P. 179-190. 15. Lu С., Vonshak A. Characterization of PSII photochemistry in salt-adapted cells of cyanobacterium Spirulina platensis // New Phytologist. 1999. V.141. P. 231-239. 17. Peloquin J.A., Smith W.O. Jr. The role of phytoplankton size on photochemical recovery during the Southern Ocean Experiment // J. Phycol. 2006. V. 42. P. 1016–1027. 18. Reeves S., McMinn A., Martin A. The effect of prolonged darkness on the growth, recovery and survival of Antarctic sea ice diatoms // Polar Biol. 2011. V. 34. P. 1019–1032. 19. Rijstenbil J. W. UV- and salinity-induced oxidative effects in the marine diatom Cylindrotheca closterium during simulated emersion // Mar. Biol. 2005. V. 147. P. 1063–1073. 20. Rijstenbil J.W., Mur L.R., Wijnholds J.A., Sinke J.J. Impact of a temporal salinity decrease on growth and nitrogen metabolism of the marine diatom Skeletonema costatum in continuous cultures // Mar. Biol. 1989a. V. 101. P. 121–129. 21. Rijstenbil J.W., Wijnholds J.A., Sinke J.J. Implications of salinity fluctuations for growth and nitrogen metabolism of the marine diatom Ditylum brightwellii in comparison with Skeletonema costatum // Mar. Biol. 1989b. V. 101. P.131–141. 22. Rynearson T.A., Armbrust E.V. Genetic differentiation among populations of the planktonic marine diatom Ditylum brightwellii (Bacillariophyceae) // J. Phycol. 2004. V. 40. P. 34–43. 23. Rynearson T.A., Armbrust E.V. Maintenance of clonal diversity during a spring bloom of the centric diatom Ditylum brightwellii // Molecular Ecology. 2005. V. 14. P. 1631-1640. 24. Shulaev V., Cortes D., Miller G., Mittler R. Metabolomics for plant stress response // Physiologia plantarum. 2008. V. 132. P. 1998-2008. 25. Stachura-Suphoples K., Williams D.M. Description of Conticribra tricircularis, a new genus and species of Thalassiosirales, with a discussion on its relationship to other continuous cribra species of Thalassiosira Cleve (Bacillariophyta) and its freshwater origin // Eur. J. Phycol. 2009. V. 44. P.477-486. 26. Steemann Nielsen E. The use of radioactive carbon (C14) for measuring organic production in the sea // J. Cons. per. int. l’exporation de la mer. 1952. V. 18. P. 117-140. 27. Veldhuis M.J.W., Kraay G.W., Timmermans K.R. Cell death in phytoplankton: correlation between changes in membrane permeability, photosynthetic activity, pigmentation and growth // Eur. J. Phycology. 2001. V. 36. P. 167–177. опубликовано - декабрь 2012 г.
Fluorescence of individual cells of the algae Conticribra weissflogii under hyperosmotic stress Effects of salinity increase to 35 psu on growth, photosynthetic carbon fixation, relative variable fluorescence of individual cells Fvi/Fmi of diatom Conticribra weissflogii acclimated to salinity 17,5 psu were studied. A significant heterogeneity of the population in terms of Fvi/Fmi was revealed. Osmotic stress resulted in an increase in the variability of Fvi/Fmi. Key words: Fluorescence; population heterogeneity; osmotic stress; Conticribra weissflogii.
Об авторах Ильяш Людмила Васильевна - Ilyash Ludmila Vasil'evna доктор биологических наук, профессор
Ilyashl@mail.ru Курочкина Виктория Александровна - Kurochkina Viktoria Alexandrovna аспирант ФГБОУ ВПО «Московский государственный университет имени М.В. Ломоносова» (Lomonosov Moscow State University, Moscow, Russia), Биологический факультет, кафедра гидробиологии vika230307@ya.ru Белевич Татьяна Алексеевна - Belevich Tatiana Alekseevna кандидат биологических наук 3438083@list.ru Погосян Сергей Иосифович - Pogosyan Sergei Iosifovich доктор биологических наук, профессор pogosyan@biophys.msu.ru Корреспондентский адрес: 119991, Россия, Москва, Ленинские горы, д. 1, стр. 12, Биологический факультет МГУ, каф. гидробиологии; телефон (495) 939-11-48
ССЫЛКА НА СТАТЬЮ: Ильяш Л.В., Курочкина В.А., Белевич Т.А., Погосян С.И. Флуоресценция отдельных клеток водоросли Conticribra weissflogii при гиперосмотическом стрессе // Вопросы современной альгологии. 2012. № 2 (2). URL: http://algology.ru/131

|
|||
| |||